



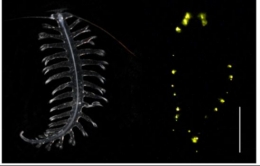

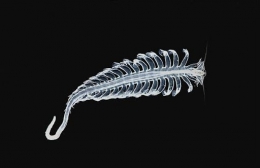
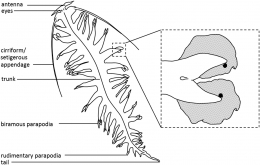

Pernahkah kamu melihat cacing laut? Berbeda dari cacing tanah yang mudah ditemukan di daratan, cacing laut agaknya sulit ditemukan karena hewan ini terletak di zona laut terdalam (deep sea). Akan tetapi, meskipun cacing laut terletak di dalam laut yang gelap, ternyata cacing laut memiliki kemampuan untuk bercahaya sehingga membuatnya menjadi organisme laut yang cukup kontras dan mudah terlihat. Salah satu spesies cacing laut yang memiliki kemampuan tersebut adalah Tomopteris helgolandica. Yuk, kita kenali jenis cacing laut yang satu ini!
Berikut adalah klasifikasi dari spesies Tomopteris helgolandica :
Kingdom: Animalia
Filum: Annelida
Kelas: Polychaeta
Ordo: Phyllodocida
Superfamili: Typhloscolecoidea
Famili: Tomopteridae
Genus: Tomopteris
Spesies: Tomopteris helgolandica
Tomopteris helgolandica merupakan spesies cacing laut yang memiliki bentuk tubuh relatif pendek dan lebar (umumnya berukuran 20 -- 100 mm), memiliki 34 segmen, memiliki ekor yang berukuran sepertiga dari total panjang tubuh, serta memiliki parapodia. Parapodia yang dimiliki oleh cacing ini berukuran panjang, dilengkapi dengan organ roset di ujung kedua cabang, dan tidak memiliki chaetae.
Ciri tubuh lain yang dimiliki oleh cacing ini adalah memiliki prostomium dengan dua antena pendek divergen, dua mata dengan lensa, dan dua organ nuchal bersilia. Spesies ini biasanya berwarna lurik dengan garis-garis coklat tua, kuning, dan atau berwarna transparan. Cacing laut yang merupakan organisme planktonik dapat mencapai tembus pandang yang hampir sempurna melalui transparansi, sehingga menjadikannya salah satu adaptasi yang paling berharga dan menarik di lingkungan pelagis.
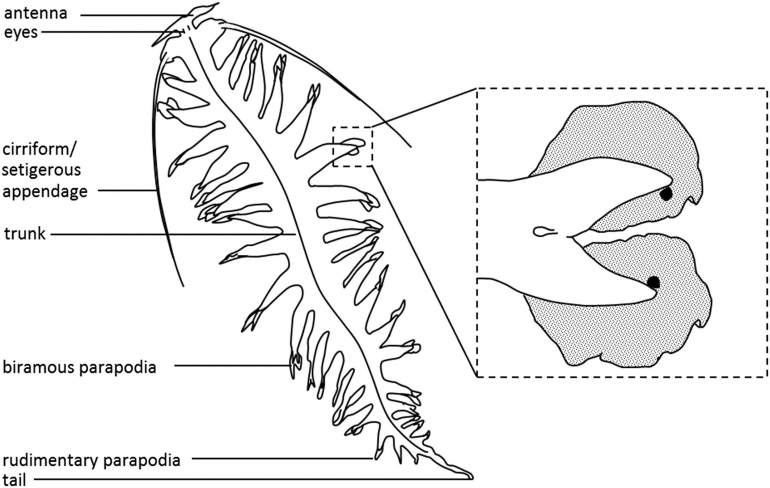
Tomopteris helgolandica memiliki habitat di perairan laut dalam (deep sea). Sebagai kelompok holoplanktonik, cacing ini persebarannya dimulai dari permukaan air sampai ke kedalaman 5000 m. Cacing juvenile cukup sering ditemukan di lapisan permukaan air, akan tetapi jarang ditemukan di lapisan permukaan air saat sudah dewasa. Hal ini disebabkan karena cacing dewasa hanya dapat ditangkap di dekat bagian bawah pada kedalaman minimal 60 meter atau lebih.
Selain itu, pengamatan juga menunjukkan bahwa pemijahan mungkin terjadi di dekat permukaan air. Spesies ini tersebar di seluruh Laut Utara, termasuk Laut Wadden, Skagerrak, Kattegat, hingga Teluk Kiel. Selain itu, spesies ini juga dapat ditemukan di Samudera Arktik, Samudera Atlantik, Selat Inggris, dan Laut Mediterania.

Anggota kelas Polychaeta kebanyakan bereproduksi secara seksual. Tahapan reproduksi ini dimulai dengan cacing betina menghasilkan feromon yang bertujuan untuk menarik dan memberikan sinyal kepada cacing jantan untuk mengeluarkan sperma, sehingga nantinya akan merangsang cacing betina untuk melepaskan telur. Perilaku ini dikenal sebagai swarming.
Setelah melewati tahap tersebut, nantinya gamet akan dilahirkan melalui metanephridia atau dinding tubuh yang pecah. Setelah pembuahan, sebagian besar telur akan berkembang menjadi plankton, meskipun ada juga yang tertahan di dalam tabung atau terkubur di dalam massa jeli yang menempel pada tabung penetas telur. Siklus hidup yang akan dilalui oleh cacing laut adalah telur berkembang menjadi larva trocophore, kemudian bermetamorfosis menjadi cacing juvenile (tubuh memanjang), dan terakhir akan berkembang menjadi cacing dewasa.
Tomopteris helgolandica merupakan spesies polychaeta holoplanktonik transparan yang memiliki kemampuan untuk menghasilkan pendaran cahaya bioluminescence. Meskipun terdapat beberapa organisme yang menghasilkan cahaya bioluminescence secara terus menerus, ternyata sebagian besar sinyal emisi cahaya adalah peristiwa sementara yang dimediasi oleh mekanisme kontrol khusus.
Mekanisme kontrol khusus terdiri dari kontrol ekstrinsik yang diwakili oleh jalur kontrol perifer, serta kontrol intrinsik yang mencakup reaksi fotogenik dan jalur pensinyalan intraseluler. Dalam metazoan self-luminescent yang dicirikan oleh struktur fotogenik yang berbeda, emisi cahaya dikendalikan oleh hormon atau melalui mekanisme penggabungan antara fotosit dan sel yang dapat dirangsang, termasuk sel saraf, sel otot, atau sel epitel.
Akan tetapi, sesungguhnya pendaran cahaya paling sering dikendalikan oleh jalur saraf. Selain berfungsi untuk menyalakan dan mematikan emisi cahaya, kemampuan kontrol saraf dapat memodulasi dan menyesuaikan intensitas, durasi, frekuensi, atau distribusi sudut dari sinyal cahaya, serta dapat menghasilkan keragaman dan spesifisitas. Namun, sebagian besar keragaman fungsional dari pola emisi dan sistem kontrol masih belum diketahui, terutama pada Annelida, di mana studi kontrol bioluminesensi masih terbatas pada spesies bentik polinoid dan chaetopterid. Maka dari itu, kontrol pendaran spesies cacing pelagis ini masih perlu untuk diteliti lebih lanjut.

Terlepas dari perbedaan yang diamati dari pola emisi cahaya, kontrol, dan karakteristik bioluminesensi antara cacing polinoid dan chaetopterid, banyak kasus yang telah mengaitkan bioluminesensi dengan fungsi pertahanan. Beberapa jenis cacing laut menggunakan emisi cahaya mereka sebagai upaya pencegahan dan komunikasi intraspesifik selama masa kawin.
Mengingat bahwa laut tidak memfasilitasi kontak antara organisme planktonik, ternyata panjang gelombang emisi atipikal akan sangat menguntungkan bagi kelompok Tomopteridae. Meskipun pancaran cahaya kuning telah ditafsirkan oleh banyak peneliti sebagai sinyal spesifik yang melibatkan saluran komunikasi pribadi, namun demikian, ada kemungkinan bahwa cahaya kuning yang dihasilkan oleh Tomopteris helgolandica memainkan peranan yang berbeda. Hal ini menunjukkan bahwa cacing laut memodulasi output cahaya dengan bergantung pada konteks yang menghasut penggunaan bioluminesensi.
Sekian penjelasan mengenai spesies cacing laut Tomopteris helgolandica. Semoga artikel ini dapat bermanfaat bagi kalian semua. Terima kasih!
DAFTAR ACUAN
Akesson, B. 1962. The Embryology of Tomopteris helgolandica (Polychaeta). Acta Zoologica 43(2-3): 135--199.
Anctil, M. 1987. Neural control mechanisms in bioluminescence. In Nervous Systems in Invertebrates (ed. M. A. Ali), pp. 573--602. New York, NY: Plenum Press.
Case, J.F. & L.G. Strause. 1978. Neurally controlled luminescent systems. In Bioluminescence in Action (ed. P. J. Herring), pp. 331--366. London; New York, NY; San Francisco, CA: Academic Press.
Claes, J.M. & J. Mallefet. 2009. Hormonal control of luminescence from lantern shark (Etmopterus spinax) photophores. J. Exp. Biol. 212: 3684--3692.
Couwelaar, M.V. 2003. Tomopteris helgolandica. 1 hlm. http://species-identification.org/species.php?species_group=zmns&menuentry=soorten&id=256&tab=beschrijving. 23 Desember 2021. pk. 16.38 WIB.
Dales, R.P. 1971. Bioluminescence in pelagic polychaetes. J. Fish. Res. Board Can. 28: 1487--1489.
Deheyn, D.D. & M.I. Latz. 2009. Internal and secreted bioluminescence of the marine polychaete Odontosyllis phosphorea (Syllidae). Invertebr. Biol. 128: 31--45.
Fischer, A. & U. Fischer. 1995. On the life-style and life-cycle of the luminescent polychaete Odontosyllis enopla (Annelida: Polychaeta). Invertebr. Biol. 114: 236--247.
Gardner, C.R. & R.J. Walker. 1982. The roles of putative neurotransmitters and neuromodulators in annelids and related invertebrates. Prog. Neurobiol. 18: 81--120.
Gaston, G.R. & J. Hall. 2000. Lunar periodicity and bioluminescence of swarming Odontosyllis luminosa (Polychaeta: Syllidae) in Belize. Gulf Car. Res. 12: 47--51.
Gouveneaux, A., M.-C. Gielen & J. Mallefet. 2018. Behavioural responses of the yellow emitting annelid Tomopteris helgolandica to photic stimuli. Luminescence 33(3): 511--520.
Harvey, E.N. 1952. Bioluminescence. New York, NY: Academic Press.
Hastings, J.W. & J.G. Morin. 1991. Bioluminescence. In Neural and Integrative Animal Physiology, Vol. 3 (ed. C. L. Prosser), pp. 131--170. New York, NY: Wiley-Liss.
Herring, P.J. & J.G. Morin. 1978. Bioluminescence in fishes. In Bioluminescence in Action, pp. 273--329. New York, NY: Academic Press.
Johnsen, S. & E.A. Widder. 1998. Transparency and visibility of gelatinous zooplankton from the northwestern Atlantic and Gulf of Mexico. Biology Bulletin 195: 337--348.
Krnstrm, J., W. Karlsson, B.R. Johansson & S. Holmgren. 2009. Involvement of contractile elements in control of bioluminescence in Northern krill, Meganyctiphanes norvegica (M. Sars). Cell Tissue Res. 336: 299--308.
Latz, M.I., T.M. Frank & J.F. Case. 1988. Spectral composition of bioluminescence of epipelagic organisms from the Sargasso Sea. Mar. Biol. 98: 441--446.
Nicol, J.A.C. 1960. The regulation of light emission in animals. Biol. Rev. Camb. Philos. Soc. 35: 1--40.
Salazar-Vallejo, S.I. 1996. Lista de species y bibliografa de Poliquetos (Polychaeta) del Gran Caribe. Anales Inst. Biol. Univ. nac. Autn, Mxico, Ser. Zool. 67(1): 11--50.
Tsuji, F.I. & E. Hill. 1983. Repetitive cycles of bioluminescence and spawning in the polychaete, Odontosyllis phosphorea. Biol. Bull. 165: 444--449.
Wilkens, L.A. & J.J. Wolken. 1981. Electroretinograms from Odontosyllis enopla (Polychaeta; Syllidae): initial observations on the visual system of the bioluminescent fireworm of Bermuda. Mar. Freshw. Behav. Physiol. 8: 55--66.
Baca konten-konten menarik Kompasiana langsung dari smartphone kamu. Follow channel WhatsApp Kompasiana sekarang di sini: https://whatsapp.com/channel/0029VaYjYaL4Spk7WflFYJ2H
