




Sel adalah unit struktural dan fungsional serta hereditas (pewarisan sifat) terkecil penyusun makhluk hidup dalam tingkatan-tingkatan organisasi kehidupan. Kata "sel" berasal dari bahasa yunani, yaitu "cellula" atau "cella" yang berarti ruang kosong. Pengertian pertama tentang sel ini dikemukakan oleh Robert Hooke. Robert Hooke meneliti sel gabus yang merupakan sel tumbuhan mati. Dari sinilah bermula semua penelitian tentang sel. Sel menjadi penyusun dasar setiap tubuh makhluk hidup. Sel mampu melakukan semua aktivitas kehidupan dan sebagian besar dari reaksi kimia untuk mempertahankan kehidupan berlangsung di dalam sel.
Kebanyakan makhluk hidup tersusun atas sel tunggal atau organisme uniseluler, misalnya bakteri dan amoeba. Makhluk hidup lainnya, termasuk tumbuhan, hewan, dan juga manusia, merupakan organisme multiseluler yang terdiri dari banyak tipe sel yang terspesialisasi dengan fungsinya masing-masing. Tubuh manusia, misalnya, tersusun atas lebih dari 1013 sel. Namun, seluruh tubuh semua organisme berasal dari hasil pembelahan suatu satu sel.
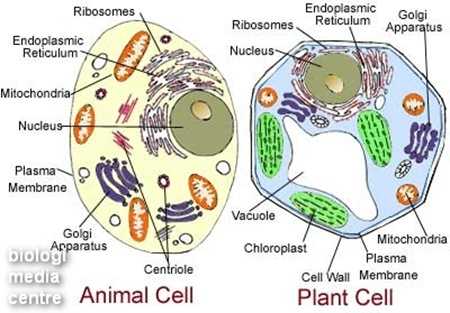
Sel-sel pada organisme multiseluler tidak akan bertahan lama jika masing-masing berdiri sendiri. Sel yang sama dikelompokkan menjadi jaringan, yang membangun organ dan kemudian sistem organ yang membentuk tubuh organisme tersebut.Di dalam sel terdapat berbagai organel sel yang memiliki bentuk, struktur, dan fungsi yang khas. Masing-masing bekerja sama untuk menjaga kelangsungan hidup sel. Salah satu organel terpenting yang berperan dalam penyuplai energi adalah mitokondria.
Mitokondria adalah tempat di mana fungsi respirasi pada makhluk hidup berlangsung. Respirasi merupakan proses perombakan atau katabolisme untuk menghasilkan energi atau tenaga bagi berlangsungnya proses hidup. Dengan demikian, mitokondria adalah pembangkit tenaga bagi sel. (Purnobasuki, 2011)
Mitokondria menyuplai energi dengan berbagai macam cara meperti melakukan metabolismeasam lemak, biosintesispirimidina, homeostasiskalsium, dan transduksi sinyal seluler. Metabolisme dalam makhluk hidup dapat dibedakan menjadi dua, yaitu katabolisme dan anabolisme. Secara singkat, katabolisme adalah pemecahan senyawa menjadi lebih sederhana. Katabolisme menghasilkan energi. Sedangkan anabolisme sebaliknya. Metabolisme asam lemak adalah pemecahan senyawa lemak untuk mendapatkan energi.
Selain itu, fungsi lain dari mitokondria adalah sel signal untuk neuron, mengelola apoptosis, mengontrol siklus sel dan pemantauan diferensiasi sel pertumbuhan dan perkembangan. Apoptosis adalah mekanisme biologi multi sel untuk membuang sel yang sudah tidak diperlukan lagi. Di sini peran mitokondria adalah menghentikan suplai energi sehingga sel tersebut mati dan tidak bertumbuh menjadi sel pengganggu. Peran mitokondria dalam pertumbuhan dan perkembangan sel adalah mengontrol seberapa banyak energi yang diperlukan sel yang sedang mengalami perkembangan.
Berhubung dengan fungsi respirasi mitokondria, mitokondria banyak terdapat pada sel yang memilki aktivitas metabolisme tinggi dan memerlukan banyak ATP dalam jumlah banyak, misalnya sel otot jantung. Jumlah dan bentuk mitokondria bisa berbeda-beda untuk setiap sel. Pada umumnya, semakin sedikit jumlah mitokondria dalam suatu sel, maka semakin besar ukuran organel mitokondria. Misal pada spesies ganggang yang tidak berwarna tidak mempunyai mitokondria. Spermatozoa dan flagellata tertentu hanya memiliki satu mitokondria per sel, hati memiliki kurang lebih 800 mitokondria per sel. Pada beberapa keadaan terdapat kaitan langsung antara jumlah mitokondria per sel dengan keperluan metabolisme sel.
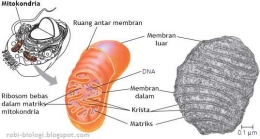
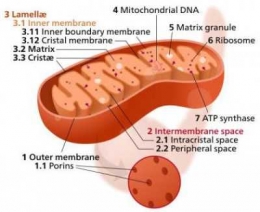
"Mitokondria berbentuk elips dengan diameter 0,5 m -- 1,0 m dan panjang 1 -3 m. Struktur mitokondria terdiri dari empat bagian utama, yaitu membran luar, membran dalam, ruang antar membran, dan matriks yang terletak di bagian dalam membran." (Cooper, 2000)
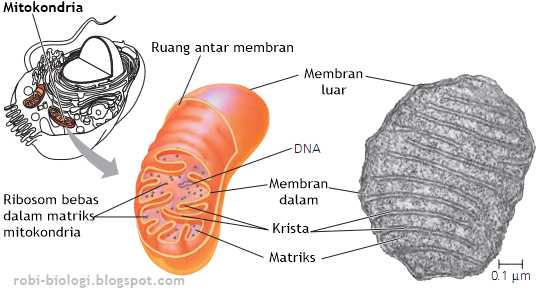
Dengan sifat membran yang permiabel terhadap molekul molekul di sitosol, maka pada ruang antar membran secara kimiawi berisi cairan yang sama seperti di sitosol. Ruang antar membarn mengandung beberapa enzim yang digunakan untuk mengeluarkan ATP dari matrix untuk memfosfolirasi nuleotida lain ( Albert, 2008 : 818)
Membran dalam bersifat kurang permeabel dibandingkan membran luar terdiri dari 20% lipid dan 80% protein. Membran ini merupakan tempat utama pembentukan ATP (Adenosin Tri Phosphat). Luas permukaan ini meningkat sangat tinggi karena banyaknya lipatan yang menonjol ke dalam matriks, disebut Krista. Stuktur krista berfungsi untuk meningkatkan luas permukaan membran dalam sehingga meningkatkan kemampuannya dalam memproduksi ATP. Membran dalam mengandung protein yang terlibat dalam reaksi fosforilasi oksidatif, ATP sintase yang berfungsi membentuk ATP pada matriks mitokondria, serta protein transpor yang mengatur keluar masuknya metabolit dari matriks melewati membran dalam.
Struktur morfologi yang paling bervariasi dalam mitokondria adalah krista. Dalam satu sel tertentu krista biasanya seragam dan khas bagi sel itu. Dalam tipe-tipe sel yang berbeda, bentuk krista bisa sangat berbeda. Sebagian besar mitokondria mempunyai krista yang berbentuk seperti lamela atau seperti tubul (tabung). Krista yang berbentuk seperti lamela adalah jenis krista yang paling umum, lamela relatif paralel atau bertumpuk-tumpuk teratur.

Mitokondria memiliki keunikan tersendiri dari organel lain di dalam sel. Mitokondria merupakan organel yang mengandung DNA, selain DNA inti pada nukleus. Dari keunikan inilah para peneliti berusaha mencari informasi lebih lanjut, peristiwa ini mempelopori perkembangan penelitian mitokondria.
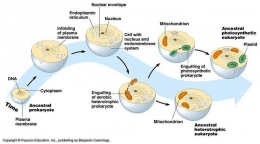
Pada tahun 1981 Lynn Margulis menciptakan Teori Endosimbiosis Serial (TES), yaitu evolusi sel eukariot melibatkan simbiosis dari beberapa sel nenek moyang yang saling bebas dalam urutan yang spesifik. Para nenek moyang ini terdiri atas sel inang (arkea metanogen serupa Thermoplasma), nenek moyang mitokondria (serupa Daptobacter dan Bdellovibrio), nenek moyang kloroplas (serupa Cyanobacter) dan nenek moyang struktur pergerakan selular (serupa Spirochete). Para nenek moyang simbion masuk ke sel inang sebagai makanan yang tidak dicerna atau sebagai parasit internal yang kemudian antar mereka saling bekerjasama yang disebut endosimbiosis. Ketika mereka menjadi saling tergantung maka simbiosis antar mereka menjadi saling tak terpisahkan. (Marguilis, 1981)

Berdasarkan hipotesis endosimbiosis mitokondria berasal dari sel eukariotik yang bersimbiosis dengan prokariot (bakteri) sehingga membentuk organel sel. Adanya DNA pada mitokondria menunjukkan bahwa dahulu mitokondria merupakan entitas yang terpisah dari sel inangnya. Hipotesis ini ditunjang oleh beberapa kemiripan mitokondria dengan bakteri. Mitokondria ini menyerupai bakteri mulai dari bereproduksi dengan cara membelah diri menjadi dua, memiliki sistem genetik sendiri, dan memiliki ribosom. Ribosom mitokondria lebih mirip dengan bakteri dibandingkan dengan ribosom yang dikode oleh inti sel eukariot.
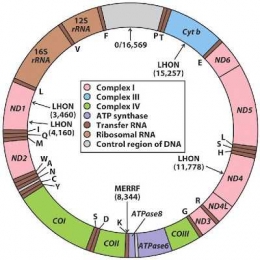
Asal-usul mitokondria sebagai entitas yang terpisah maka mitokondria memiliki DNA sendiri. DNA mitokondria berbeda namun berhubungan dengan DNA yang dimiliki nucleus. DNA mitokondria terdapat dalam bentuk sirkular tunggal atau "catenated", yaitu dua atau lebih untai berkaitan bersama sama seperti kaitan dalam rantai, didalam matriks mitokondria. Satu mitokondria biasanya memiliki 2-6 kopi DNA, sehingga jumlah mtDNA per sel mencapai 108atau bergantung pada jumlah mitokondria tipe tertentu (Siregar,1990).

Sistem semiotonom mitokondria terlihat dari cara sintesis ribosom mitokondria. RNA ribosom mitokondria ditanskripsi di mtDNA sedang protein ribosom mitokondria ditranskripsi dari DNA inti, kemudian ditranslasi pada ribosom sitoplasma dan akhirnya diangkut ke dalam mitokondria untuk perakitan partikel nukleotida (ribosom) (Siregar, 1990).
Mitokondria memiliki sifat untuk memperbanyak diri. Peristiwa ini disebut dengan mitokondriogenesis. Proses pembantukan mitokondria terdiri dari dua tahap, yakni tahap pertumbuhan dan reproduksi. Proses diferensiasi organel tersebut tak lepas dari peran DNA mitokondria. Proses diawali dengan pemanjangan mitokondria, selanjutnya satu atau lebih krista yang berada ditengah akan membentuk ruangan dengan tumbuh memotong matriks dan melebur dengan membran dalam di sampingnya. Setelah bagian dalam terbelah menjadi dua, membran luar mengalami invaginasi (pemisahan) yang dilanjutkan dengan peleburan kedua membran dalam. Pada akhirnya terbentuklah dua mitokondria.
Setiap individu tumbuh dan berkembang selalu bermula dari peleburan dua sel (sel ovum dan sel sperma). Keduanya, sel sperma dan sel ovum masing-masing memiliki organel sel yang sama. Masing-masing sel telur dan sel sperma pasti memiliki nukleus (intisel), sitoplasma, retikulum endoplasma, badan golgi, ribosom, lisosom, mitokondria, dan berbagai organel sel lain. Namun, dalam penelitian beberapa ahli menyatakan bahwa mitokondria yang ada dan berfungsi di dalam tubuh kita merupakan mitokondria yang berasal dari sel ovum saja, tidak ada mitokondria yang berasal dari sel sperma. Pertanyaan utamanya adalah mengapa mitokondria di tubuh kita tidak berasal dari kedua sel orang tua kita.
Dalam proses pembuahan, jutaan sel sperma akan melintasi rongga rahim, saling berebut untuk mencapai sel telur matang yang berada pada saluran oviduk (saluran oviduk atau fuba falopi adalah saluran kecil yang menghubungkan uterus dengan kantung telur) di seberang rahim. Dari jutaan sel sperma hanya akan ada satu sel sperma yang masuk ke dalam sel sperma. Meski kadang-kadang terjadi anomali, ada dua sel sperma yang masuk ke dalam sel ovum (keadaan ini menyebabkan kembar tidak identik). Setelah satu sel sperma berhasil masuk ke dalam sel ovum, maka akan terjadi perubahan-perubahan pada permukaan sel telur dan tidak dapat dimasuki lagi oleh sperma. Berjuta-juta sel sperma lain akan mati dengan sendirinya.
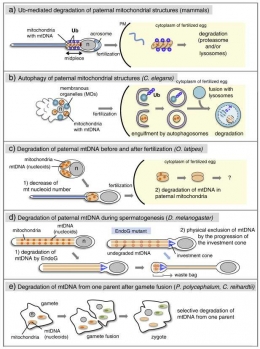
Ketika sel telur dibuahi, di dalam inti sel telur akan terjadi reaksi persenyawaan antara sifat-sifat atau kromosom-kromosom dari sel telur dan juga sel sperma. Seluruh badan sel sperma masuk ke dalam ovum. Kedua mitokondria dari sel ayah (paternal mitokondria) dan ibu (maternal mitokondria) bertemu saat pembuahan, namun biasanya paternal mitokondria menghilang saat tahap awal embriogenesis. Alasan dari mekanisme pertahanan tubuh manusia yang menghilangkan paternal mitokondria belum diketahui secara pasti.
Penelitian terbaru, menunjukkan bahwa lisosom berperan dalam mem"autofag" paternal mitokondria dalam embrio Caenorhabditis elegans. Setelah terjadi fertilisasi, lisosom berkumpul mengelilingi sel sperma yang berhasil masuk dalam sel telur. Kemudian lisosom akan menelan paternal mitokondria saat tahap awal embriogenesis.
Dalam kebanyakan spesies termasuk manusia, mtDNA yang diwariskan mitokondria merupakan sepenuhnya, murni dari sel telur. Pola pewarisan sifat ini yang dikenal dengan sebutan "maternal inheritance of mtDNA" sudah menjadi hal yang pokok untuk diketahui dalam evolusi manusia. Sama seperti mamalia lain, paternal mitokondria masuk ke dalam sel telur melalui pembuahan, tapi akan menghilang saat tahap perkembangan embrio ke-16. Setelah sel sperma masuk, lisosom yang melayang-layang di sitoplasma akan berkumpul di sekitar sel sperma dan menandai paternal mitokondria serta mulai men"autofag" atau menelan.
Dari seluruh populasi mitokondria, lisosom dapat memilah secara tepat dan hanya menelan paternal mitokondria. Lisosom dapat memilah secara tepat karena ada signal dari suatu senyawa yang dikandung oleh paternal mitokondria. Dari penelitian Miyuki Sato dan Ken Sato, paternal mitokondria sudah ditandai dengan protein ubiquitin sebelum pembuahan. Paternal mitokondria yang membawa protein ubiquitin akan langsung diserang dan ditelan oleh lisosom. Selain itu, signal kuat yang dipancarkan oleh protein ubiquitin bukan hanya dari paternal mitokondria tetapi juga ditemukan di organel bermembran (Membranous Organelles). MOs juga diasingkan dari embrio oleh lisosom. Benang merah yang diketahui dari penelitian ini adalah paternal mitokondria dikenali oleh lisosom berdasarkan protein ubuquitin yang dikandung oleh organel bermembran.
Di samping itu, pengenalan paternal mitokondria juga dapat diketahui dari karakteristik morfologi paternal mitokondria berbentuk granular. Bentuk granular ini yang dapat membedakan dari mitokondria lain.
"Paternal mitochondria also seem to be proliferation- and fusion-inactive, because those inherited in the lgg-1 mutant retain the granular morphology and do not increase in number during embryogenesis." (www.landesbioscience.com, diakses tanggal 4 Agustus 2017)
Paternal mitokondria secara kimia menjadi tidak aktif dan tidak dapat melakukan fungsinya, sebagai penyuplai energi, kemungkinan karena bentuk morfologinya. Paternal mitokondria juga tidak melakukan pembelahan diri saat awal tahap embriogenesis. Maka lisosom menganggap paternal mitokondria sebagai benda pengganggu kerja tubuh embrio. Benda asing ini kemudian ditelan oleh lisosom agar tidak menimbulkan efek lain dalam pertumbuhan dan perkembangan embrio. Dengan kata lain, sebelum pembuahan terjadi paternal mitokondria sudah terprogram untuk mengurangi jumlahnya secara cepat di dalam embrio tersebut. Hal ini dibuktikan dengan jumlah mtDNA pada sitoplasma sel sperma di tahap awal spermatid berjumlah 50 mtDNA, namun jumlah mtDNA berkurang drastis menjadi 10 mtDNA saat proses menghasilkan sel sperma yang matang.
"Another important question is why paternal mitochondria and mtDNA should be eliminated from embryos. One hypothesis is that the paternal mitochondria and/or mtDNA are heavily damaged by reactive oxygen species prior to fertilization, and are hence removed to prevent the spread of potentially deleterious mitochondria to the whole population."(www.landesbioscience.com, diakses tanggal 4 Agustus 2017)
Banyak ahli berusaha menyimpulkan mengapa paternal mitokondria harus hilang dalam proses pertumbuhan dan perkembangan janin. Salah satu hipotesis mengatakan bahwa paternal mitokondria dan/atau mtDNA akan bereaksi dengan suatu jenis oksigen dan mengakibakan kerusakan. Maka karena paternal mitokondria dan/atau mtDNA mengalami kerusakan keduanya akan dihancurkan oleh lisosom untuk mencegah kerusakan pada mitokondria yang lain diseluruh jaringan embrio.
Hipotesis lain mengatakan bahwa tujuan penghilangan paternal mitokondria untuk mencegah terjadinya pencampuran mitokondria yang menyebabkan mutasi. Karena mitokondria adalah organel khusus yang membawa materi genenetik (DNA) sendiri, maka pencampuran dan mutasi DNA tersebut dapat terjadi bila kedua mitokondria ada dalam satu tubuh. Bila terjadi proses pencampuran gen, mitokondria bisa saja mereplikasi secara cepat karena kehilangan gen yang mengontrol pembelahan mitokondria. Bila jumlah mitokondria terlalu banyak, maka produksi energi bagi jaringan tersebut tidak akan efisien.

Maka kesimpulan yang dapat ditarik yaitu pertumbuhan dan perkembangan embrio yang menjadi suatu individu secara normalnya hanya mengandung maternal mitokondria. Pewarisan mitokondria tunggal dari orang tua, memiliki proses yang berbeda-beda tergantung dari spesies masing-masing. Dapat disimpulkan bahwa proses penghilangan paternal mitokondria dilakukan dengan berbagai cara, antara lain: pengurangan jumlah mtDNA mitokondria pada proses spermatogenesis, mengeliminasi mtDNA dari proses pematangan sperma, pencegahan masuknya paternal mitokondria ke dalam sel ovum dengan cara melepas sebagian ekor sel sperma, dan pemrograman paternal mitokondria secara alami untuk tidak melakukan pembelahan saat embriogenesis. Selain itu, proses autofag oleh lisosom saat paternal mitokondria berhasil masuk dalam sel telur. Proses autofag ini terjadi karena paternal mitokondria memiliki program secara alami untuk tidak melakukan pembelahan dan paternal mitokondria yang berhasil masuk tidak akan berfungsi secara efisien. Maka, paternal mitokondria dianggap sebagai benda asing dan akan ditelan oleh lisosom yang berfungsi sebagai mekanisme pertahanan sel.
DAFTAR PUSTAKA:
- Albert, Bruce el all. 2008. Molecular Biology of The Cell Fifth Edition. USA : Garland Science Taylor & Francis Group.
- Bawa, Wayan.1998. Dasar-Dasar Biologi Sel. Jakarta: Depdikbud Dikti PPLPTK
- Lodish, H. 2001. Molecular Cell Biology 4th edition. England : W.H. Freeman and Company.
- Marguilis, L. 1981. Symbiosis in Cell Evolution. England : W.H. Freeman and Company.
- M. Sato, K. Sato.2013. Maternal inheritance of mitochondrial DNA by diverse mechanisms to eliminate paternal mitochondrial DNA, Biochim. Biophys. Acta
- Siregar, Arbayah. 1990. Biologi Sel. Bandung : FMIPA Institut Teknologi Bandung.
- http://www.lifesci.sussex.ac.uk/home/Adam_EyreWalker/Website/Publications_files/BromhamTREE03.pdf . Mitochondrial Steve: paternal inheritance of mitochondria in humans . diakses tanggal 4 Agustus 2017.
- http://www.docs-engine.com/pdf/1/Maternal-inherintance-of-human-mitochond-rial.html. diakses tanggal 4 Agustus 2017.
- https://www.researchgate.net/profile/Tri_Susmiarsih/publication/293556782_Peran_genetik_DNA_mitokondria_mtDMA_pada_motilitas_spermatozoa/links/56b98b5608ae3b658a88cf3f/Peran-genetik-DNA-mitokondria-mtDMA-pada-motilitas-spermatozoa.pdf.
- diakses tanggal 12 Agustus 2017
- https://id.wikipedia.org/wiki/Mitokondria. diakses tanggal 17 Agustus 2017
- https://id.wikipedia.org/wiki/Sel_(biologi). diakses tanggal 20 Agustus 2017
- http://www.academia.edu/5569741/TEORI_ENDOSIMBIOSIS. diakses tanggal 17 Agustus 2017
- http://www.generasibiologi.com/2016/10/struktur-dan-fungsi-organel-mitokondria.html. diakses tanggal 17 Agustus 2017
Baca konten-konten menarik Kompasiana langsung dari smartphone kamu. Follow channel WhatsApp Kompasiana sekarang di sini: https://whatsapp.com/channel/0029VaYjYaL4Spk7WflFYJ2H
